
 首页
首页 登录
登录 注册
注册


-
冷大气压等离子体(cold atmospheric plasma, CAP)由带电粒子、活性物质等组成, 由于其高度的反应活性和温和的产生条件, CAP被广泛地应用于医学领域[1–4], 例如: 伤口处理[5]、微生物灭活[6–8]、牙科[9]、皮肤病治疗[10]等. 近年来, CAP被发现具有杀伤癌细胞而不影响正常细胞的“选择性”抗癌效应[11–13]. 为此, 学界进行了大量关于CAP杀伤癌细胞的宏观和微观机制的研究, 目前的主流观点有两种: 1) CAP诱导的电场可以影响细胞膜的流动性和渗透性, 导致细胞膜发生电穿孔(electroporation, EP)从而干扰癌细胞微环境[14]; 2) CAP含有的活性物质, 包括活性氧物质(reactive oxygen species, ROS)和活性氮物质(reactive nitrogen species, RNS), 可以进入癌细胞引起氧化应激, 从而触发癌细胞的凋亡程序[15].
EP被认为是一种安全有效的新型抗癌电化学疗法, 可运输癌症治疗相关的重要物质, 例如基因电转移、药物递送和CAP所产生ROS的运输[16–18]. Szlasa等[19]研究发现对A375细胞施加脉冲电场后, 细胞中的ROS含量显著增加, 由此提出一种假设: 电极附近溶液电解产生的ROS在电场作用下渗透细胞膜, 并进入细胞内部引发凋亡, 但对其微观机理并没有给出解释. 目前对于CAP引发细胞膜EP的研究往往围绕着EP本身的过程开展, 忽视了探讨EP形成的亲水孔在转运ROS中的作用和潜力. 因此, 从微观角度详细地解释EP形成过程, 以及根据不同的输送物质选择适合的CAP治疗参数仍然是该方法实际应用面临的关键挑战.
近年来, 随着计算机性能的迭代增强以及分子模拟技术的发展, 分子动力学模拟(molecular dynamics, MD)方法已经可以在较大体系和时间尺度下进行生理过程的模拟[20–23], 并且可以研究多种细胞在原生环境和外部干预下生理活动的微观分子机制(如神经信号传递[24]、靶向药物结合[25]、免疫抗原抗体结合[26]等), 为实际医学应用提供指导, 这为解释CAP激发电场导致EP的微观机理和分子机制提供了新的手段.
本文在Gromos 54a7力场下模拟了代表正常细胞和癌细胞膜差异的不同胆固醇含量的细胞膜发生EP的电场强度阈值、EP发生时间, 并阐明了EP进展过程的物理机制, 直观地对比了正常细胞和癌细胞对EP的抵抗性差异. 其次, 使用伞状采样法计算了不同胆固醇含量的细胞膜中, EP形成各个阶段ROS物质在其中转运过程的自由能剖面, 对EP和ROS转运的协同效应进行了深入探讨, 为CAP的“选择性”抗癌作用提供了微观解释.
-
本文选取1-棕榈酰基–2-油酰基磷脂酰胆碱(1-palmitoyl-2-oleoyl-sn-glycero-3-pc, POPC)分子和胆固醇分子作为细胞膜模型的主要成分. POPC分子是细胞生物膜的主要成分, 广泛应用于各类细胞膜建模中; 胆固醇分子在维持细胞膜结构, 提高细胞膜流动性以及扩大细胞膜适应的温度范围上有重要作用. 本文使用的POPC分子和胆固醇分子结构如图1(a)所示. 细胞膜模型用Genmixmem软件(
http://sobereva.com/245 )建立, 如图1(b)所示, 具体建模过程如下: 首先, 设定模型成分参数和盒子大小, 将预设参数输入Genmixmem软件进行POPC和胆固醇的堆积; 其次, 对生成的双层膜模型进行溶剂化; 最后, 对建立的模型体系运行等温等压系综(NPT)平衡和生成模拟, 得到模型的平衡初始构象.正常细胞中, 胆固醇可占到细胞膜分子总数的20%左右, 是细胞膜重要的组成部分, 而在癌细胞中胆固醇的含量往往较少[27,28]. 本文建立了三种细胞膜模型, 通过在模型中掺杂不同含量的胆固醇分子, 体现正常细胞和癌细胞的成分差异, 具体参数如表1所列.
图2显示了不同细胞膜模型各组分沿细胞膜法向的密度分布. 在模型1中, 水分子主要分布在上下水层和POPC分子的亲水头部区域. 随着胆固醇的增加, 疏水层逐渐增厚, 两个POPC分子层之间的距离随着胆固醇含量的增加而增加. 胆固醇分子分布于细胞膜疏水区的深处, 胆固醇区的POPC分子密度降低.
-
为了消除模型内部异常的原子堆叠和不合理的相互作用力, 同时避免过大的步长导致模拟体系崩溃, 首先对体系进行50000步步长为1 fs的基于最速下降法的能量最小化模拟, 消除非物理相互作用, 使体系势能收敛至局部极小值. 在能量优化后的低能稳态基础上, 采用2 fs步长进行100 ns的NPT系综平衡模拟, 该步长设置基于GROMOS力场推荐标准, 显著地提高运算效率的同时也保证了体系温度与压强收敛至稳态. 随后, 进行300 ns的生成模拟, 确保消除初始构象偏差[29]的同时稳定细胞膜结构, 并保证在模拟过程中不易发生结构崩溃. 在模拟中, 使用Parrinello-Rahman算法[30,31]和Nose-Hoover热浴算法[32,33]将压力和温度同时稳定到101 kPa和310 K, 温度耦合时间常数为0.5 ps, 压力耦合时间常数为5.0 ps, 压缩率为4.5×10–5 bar–1(1 bar=105 Pa); 使用POPC+胆固醇和水两个温度耦合组; 使用LINCS算法[34]约束模拟体系中所有氢原子所连接化学键的长度; 使用PME法[35]处理长程库仑力.
由于细胞膜等效电容特性, 其结构对于电场具有放大作用, 跨膜的微观电场应由跨膜电位决定[36]. 根据Hu等[37,38]的实验模型, 0.01 V/nm的全局脉冲电场可以在厚度6 nm的细胞膜上产生约2 V的跨膜电压, 对应微观电场约为0.35 V/nm. 基于此, 本文选择0.2—0.5 V/nm, 间隔0.05 V/nm的电场强度, 分别施加于生成模拟后的模型上, 确保所选电场参数覆盖CAP临床参数经细胞膜等效电容效应放大后的典型值. 每一个模型在不同电场强度下, 各重复运行10次, 每次运行10 ns生成模拟并记录水桥的连通时间, 总计运行了210次模拟, 以消除偶然误差, 并寻找不同模型发生EP的电场强度阈值.
-
研究ROS转运行为难易程度的落脚点是能量学差异, 吉布斯自由能
$ \Delta G $ 表述了由一个状态向另一个状态转化所需要跨越的能量势垒, 其值表示了状态转换的难易程度. 因此, 将ROS在整个水通道内的所处位置视为不同的状态, 计算ROS在不同位置的$ \Delta G $ 变化情况, 也可以称为自由能剖面(free energy profile, FEP), 以评估电场对ROS转运功能的影响.本文采用GROMOS 54a7力场[39]参数化的POPC和SPC水分子模型, 并采用Cordeiro研究中所提供的ROS分子的力场参数[40]和Neto和Cordeiro[41]研究中所提供的胆固醇分子的力场参数. 由于RNS受细胞膜磷脂头部基团的影响有限, 具备在完整细胞膜中转运的能力[42], 因此在本文中不再考虑. 首先对前文所建立的模型进行NPT平衡和生成模拟. 随后对每个模型在0.5 V/nm的外部电场下各运行20 ns的生成模拟, 以确保在模拟时间内观察到完整的EP过程. 在每个模型的EP过程中根据所形成水桥的宽度, 选取3种构象, 分别定义为水桥阶段、水孔阶段和通道阶段, 并选取未形成EP的初始构象作为对照, 进行ROS在EP所形成的孔状结构中转运的伞状采样模拟.
伞状采样过程如图3所示. 在模拟中沿EP定义了60个窗口, 间隔0.1 nm, 总距离为7.2 nm, 窗口的定义策略囊括了ROS分子通过EP进入细胞内的完整过程. 对于每种所选取的构象, 共进行3组采样模拟, 在每组模拟中, 选择了三种由CAP产生的代表性ROS(H2O2, HO2和OH)进行模拟. 事实上, 尽管OH在静态溶液中寿命较短(约1 ns), 在CAP的超高频脉冲电场条件下, 其在气相下的有效寿命和扩散距离超过1 ms和10 mm[43–45]; 另外, 考虑到CAP在黑色素瘤治疗中的射流直接作用方式[46], 及实验观测到的胞内OH自由基显著升高[47], 我们仍将OH纳入考虑范围. EP形成的水通道不仅为OH提供低阻力路径, 电场驱动的电泳效应进一步加速其内流. 考虑到过近的距离会导致ROS分子间的相互作用干扰采样结果, 对于每种ROS各重复12次嵌入和采样, 每次嵌入5个分子, 使采样范围大于所设定的截断半径(1.1 nm). 每次嵌入和采样结束后所有ROS分子上移0.1 nm.
考虑到在EP过程中产生的水桥极不稳定, 在采样中可能会发生结构改变, 对POPC和胆固醇分子中的所有非氢重原子施加了力常数为1000 kJ/(mol·nm)的质心位置约束, 以确保其在模拟过程中不发生结构改变. 对于每次采样, 运行3 ns的生成模拟, 其中前0.5 ns用于平衡体系, 后2.5 ns的结果被用于加权直方图分析方法的FEP计算.
-
在正常状态下, POPC分子具有亲水性的头部基团与界面水分子形成致密的氢键, 并通过氢键作用和自身高密度的空间限制作用限制水分子向膜内运动. 然而, 外电场的极化作用使得POPC头部基团的运动更加剧烈, 并可能形成缺陷, 缺陷为水分子提供了深入细胞膜的机会.
如图4所示, 细胞膜的EP过程包括三个阶段. 阶段A: 水突起的形成. 在这个阶段下, 外电场极化细胞膜, 扰乱POPC头部基团的分布, 成团的水分子在外电场和极化效应的驱动下侵入缺陷区域, 向细胞膜内侧移动. 阶段B: 水桥的形成. 细胞膜一侧的水突起与另一侧水层相连, 形成水桥. 阶段C: 通道的形成. 形成的水桥显著地降低了此处POPC尾部的疏水作用, 更多的水分子迅速沿水桥进入POPC双层内, 在极短时间内扩大水桥的半径, 自此水通道正式形成. Geboers等[48]的实验结果也显示了外加电场可诱导跨膜电位累积并引发膜结构失稳形成孔结构. 然而, 应当注意的是, 并非所有阶段A中的水突起都能形成阶段B中的水桥, 水突起处的缺陷可能会因细胞膜的自我修复效应而恢复原状, 在外电场驱动力不足的情况下(图4中0.3 V/nm), 水突起在一段时间后被挤出POPC双层外, 内侧疏水层恢复封闭. 另外, 随着电场强度的增加, EP过程发生的速度逐渐加快, 这可能与电场对POPC和水分子的极化效率高低有关.
图5从水分子和POPC总偶极矩的角度量化磷脂双分子层系统的极化过程. 在EP发生前, 水层和POPC层的有序性导致其偶极矩对外电场响应较弱. 甚至直到阶段B (水桥连通)前, 水分子和POPC的总偶极矩都没有很大变化. 而水桥形成后(阶段C), 水分子的快速极化与头部基团位移显著增加系统总偶极矩, POPC尾部的疏水作用被破坏, 进入POPC层的水分子被快速极化, POPC头部基团向POPC双层中心移动, 水分子和POPC分子沿z轴偶极矩快速增加, POPC层被完全 导通.
-
进一步分析三种细胞膜电穿孔形成时间随电场强度的变化关系, 如图6. 对于模型1而言, EP阈值在约0.3 V/nm附近, 在更低的电场强度下未发生EP. 随着胆固醇含量升高, EP阈值上升至0.35 V/nm, 且各电场强度下的EP时间与胆固醇含量呈正比. Jiang等[49]也报道了通过电子显微镜观察到电场作用下细胞膜表面形成瞬态孔结构, 以及电场强度与EP形成时间存在一定的数量关系. 由于胆固醇分子高度稳定且疏水, 其填充在POPC双层的POPC尾部区域, 有效地降低了细胞膜的流动性, 并阻止外部水分子向POPC内层的侵入. 因此, 相对癌细胞而言, 更高胆固醇含量的正常细胞发生EP的电场强度阈值更高, 更容易在CAP电场作用下保持细胞膜的选择透过性. 对于胆固醇含量约20%的正常细胞, EP阈值电场为0.35 V/nm, 发生时间约为7 ns. 此阈值与CAP治疗参数(0.3—0.5 V/nm)一致, 表明正常细胞在治疗电场下可 维持膜完整性, 而低胆固醇癌细胞(阈值0.3 V/nm)更易发生不可逆EP. 在实际应用中, 应选择合适的CAP电场参数, 一方面, 使癌细胞膜发生有效EP, 摄入更多的ROS, 引发细胞内部的氧化应激等效应, 促进癌细胞死亡; 另一方面, 避免正常细胞的细胞膜结构被破坏, 从而实现对癌细胞的杀伤而不影响正常细胞的“选择性”抗癌效果.
-
图7(a)、图8(a)及图9(a)分别为模型1, 2, 3的EP形成过程中的不同阶段构象, 并与原生细胞膜构象对比, 这些构象被用于嵌入ROS及进行伞状采样和加权直方图分析. 在模型1中, 对于原生双层细胞膜(图7(b)—(d)黑色), ROS在未发生EP的细胞膜中转运时, 能量势垒出现在POPC头部基团区域, 而在POPC尾部区域的FEP更低, 这说明由于ROS和磷脂头部基团间的氢键作用较强, 磷脂头部在限制ROS的运动中起主导作用; 而在疏水的POPC尾部区域, ROS分子仅受疏水作用的影响, FEP低于POPC头部.
随着EP进展, 头部基团向孔道内迁移并形成亲水界面, 导致尾部疏水屏障被破坏, 自由能势垒显著地降低, 从而允许亲水性ROS高效内流. 如图7(b)—(d)红色所示, 一旦水桥生成, POPC双层内生成空腔, 各种ROS的自由能势垒显著下降, ROS通过细胞膜进入细胞的障碍消失. 例如H2O2的自由能势垒相对于原生阶段降低了约8.03 kcal/mol, 在模型2和模型3中这个下降量约为7.66和7.35 kcal/mol. 可见, 胆固醇含量越低, EP对细胞膜物质渗透的促进作用越明显.
在水孔阶段, 细胞膜中心层出现了自由能势垒(图7(b)—(d)蓝色), 这个现象可能是由于POPC头部向内侧移动导致的. 水桥阶段的EP孔径较小, POPC头部基团不足以向内侧移动; 而在水通道初期阶段, EP孔径扩大, 水分子团凭借氢键作用拉动EP两侧的POPC头部基团向膜中心(即EP中心)运动, 而EP中心孔径较小, 又存在亲水基团, 因此ROS在通过中心层时受到氢键的作用, 出现了一定的自由能势垒.
随着孔径进一步扩大, POPC头部基团进一步内移, 进入水通道阶段, POPC尾部的疏水效应被头部基团隔开, 出现了自由能低谷(图7(b)—(d)绿色), 说明ROS在EP导致的头部基团连通形成的亲水孔中下可以自发运动, 通过EP进入细胞内.
当细胞膜内胆固醇含量升高时(图8和图9), ROS在不同阶段的FEP变化趋势与模型1(图7)类似, 此处不再赘述. 对于原生双层结构, H2O2分子在细胞膜两侧的自由能势垒随胆固醇含量增加逐渐升高(模型1: 10.89 kcal/mol; 模型2: 11.97 kcal/mol; 模型3: 12.56 kcal/mol). 这说明胆固醇在一定程度上增加了细胞膜的密度, 使得H2O2分子受到POPC头部基团的氢键作用更强, 更难进入细胞内. 然而对于HO2和OH分子来说, 其自由能势垒并未出现明显差异, 这可能是由于其仅有1个氢键受体(H2O2为2个), 与POPC头部基团的相互作用有限, 因此对细胞膜的密度变化并不敏感.
通过以上分析可以看出, 正常细胞与癌细胞抵抗CAP激发电场带来的EP效应存在差异. 癌细胞往往对电场更加敏感, 更容易发生EP. 以H2O2为例, 在水桥阶段, 其转运的自由能势垒分别为5.21 kcal/mol(模型3), 4.31 kcal/mol(模型2)和2.85 kcal/mol(模型1), 自由能势垒随胆固醇含量降低而降低, 表明癌细胞在EP的早期就有很大可能发生ROS的渗入, 从而更快引发细胞内的氧化应激; 更高胆固醇含量的正常细胞膜对EP的抵抗效果往往比癌细胞膜更强, 发生ROS渗透的阶段也更晚. 这一现象解释了CAP“选择性”抗癌的观点, 也证实了CAP引发的EP是ROS向癌细胞内转运的另一重要通道.
-
CAP已被广泛认可为一种安全高效的“选择性”癌症治疗方法, 但现有研究多集中于CAP引起的EP过程本身, 而较少关注由EP形成的亲水孔在转运ROS中的作用. 本文采用分子动力学方法模拟了CAP激发电场引起细胞膜EP效应的微观过程, 并建立了表示正常细胞和癌细胞成分差异的细胞膜模型, 通过不同电场强度下的多组模型的模拟, 对比讨论了不同成分细胞膜发生EP的阈值电场强度和穿孔时间差异. 此外, 研究了CAP所产生的ROS(H2O2, HO2和OH)在细胞膜EP形成的通道结构中转运的过程, 及这种过程在不同成分的细胞膜中的能量学差异. 主要结论如下.
1)细胞膜在CAP激发电场下的EP过程分为三个阶段: 水突起形成, 水桥形成和水通道形成. 电场导致的水分子和POPC分子的极化(显示为径向偶极矩的迅速升高)是EP产生和进展的重要原因.
2)细胞膜发生EP的电场强度阈值和穿孔时间均与构成细胞膜的脂质分子中的胆固醇比重成正比, 胆固醇含量越高的细胞膜EP电场强度阈值越高, EP形成的时间也越长, 显示了快速增殖的癌细胞(胆固醇含量低)相比正常细胞(胆固醇含量高)对CAP激发电场更敏感.
3)细胞膜内的胆固醇含量升高会使得H2O2通过水通道进入细胞内的自由能势垒增高, 体现在细胞膜的中心层(POPC尾部区域)密度升高; 同时胆固醇会在通道初期抵抗ROS沿水孔转运的过程, 说明正常细胞发生EP后, 细胞外ROS内流要晚于癌细胞, 因此癌细胞更容易发生氧化应激反应.
本研究表明由于正常细胞与癌细胞膜成分的不同, 使得两种细胞对EP效应的响应存在差异, 补充了对CAP“选择性”抗癌原因的解释. 同时提出了CAP激发电场产生的EP效应可以作为活性物质和抗癌药物选择性递送的潜在方式, 为CAP相关治疗手段的设计提供了新的思路.
冷大气压等离子体激发电场对细胞膜电穿孔及其转运功能的影响
Effect of cold atmospheric plasma induced electric fields on cell membrane electroporation and related transport functions
-
摘要: 冷大气压等离子体(cold atmospheric plasma, CAP)由于其具有“选择性”杀伤癌细胞的效果, 被认为是一种极具潜力的癌症治疗手段. 然而, CAP对癌细胞的“选择性”杀伤作用相关的物理模型及CAP与癌细胞相互作用的微观机理仍然匮乏. 本文采用分子动力学方法模拟了CAP激发电场引起的细胞膜电穿孔(electroporation, EP)效应, 并采用伞形采样法计算了活性氧物质(reactive oxygen species, ROS)通过EP形成的不同阶段的孔结构进入细胞内部的自由能剖面. 结果表明, 相较于正常细胞膜, 胆固醇含量较低的癌细胞膜发生EP的电场强度阈值更低, 且EP发生时间更快; 对于ROS的转运过程而言, 由于癌细胞膜胆固醇含量更低, 在EP的各个阶段下ROS的自由能势垒更低, 因此在EP的各个阶段下, ROS内流的时间均早于正常细胞. 本文从分子模拟的角度探索了CAP激发电场作用下EP的形成过程, 以及EP的不同阶段中ROS转运的潜在机会, 有利于更清楚地阐述CAP“选择性”抗癌作用的微观机理, 并为CAP癌症治疗技术、设备和手段的研发提供了重要参考, 促进了CAP在临床应用方面的发展.Abstract: Cold atmospheric plasma (CAP) is considered to be a highly promising cancer treatment method, due to its “selective” anti-cancer effect. However, the physical theoretical explanation about this effect and the microscopic interactive mechanisms between CAP and tumors are still lacking. In this work, the CAP-induced electric field-caused electroporation (EP) processes of the cell membrane are modeled based on molecular dynamics. Additionally, the umbrella sampling method is utilized to compute the free energy profile of the intracellular permeation processes of the reactive oxygen species (ROS) through EP-formed pore-like structures at different EP stages. Comparative results are shown as follows. 1) Cancer cell membranes with lower cholesterol components show lower EP-generation threshold and faster EP-formation, and 2) lower free-energy barrier and earlier occurrence of free-energy barrier reduction are shown in all EP stages in cancer cell membrane. The above results explain the difference between cancer cells and normal cells when affected by CAP. Our work delves into the formation of CAP-induced EP and the transport of ROS through EP-formed pore-like structures, which contributes to a better understanding of the microscopic mechanisms of the “selective” anti-cancer effect of CAP, and provides important references for developing CAP-based cancer treatment methods, and devices, thereby facilitating the translation of CAP into clinical applications.
-
Key words:
- cold atmospheric plasma /
- electric field /
- cell membrane electroporation /
- molecular dynamics .
-
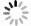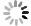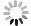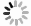
-
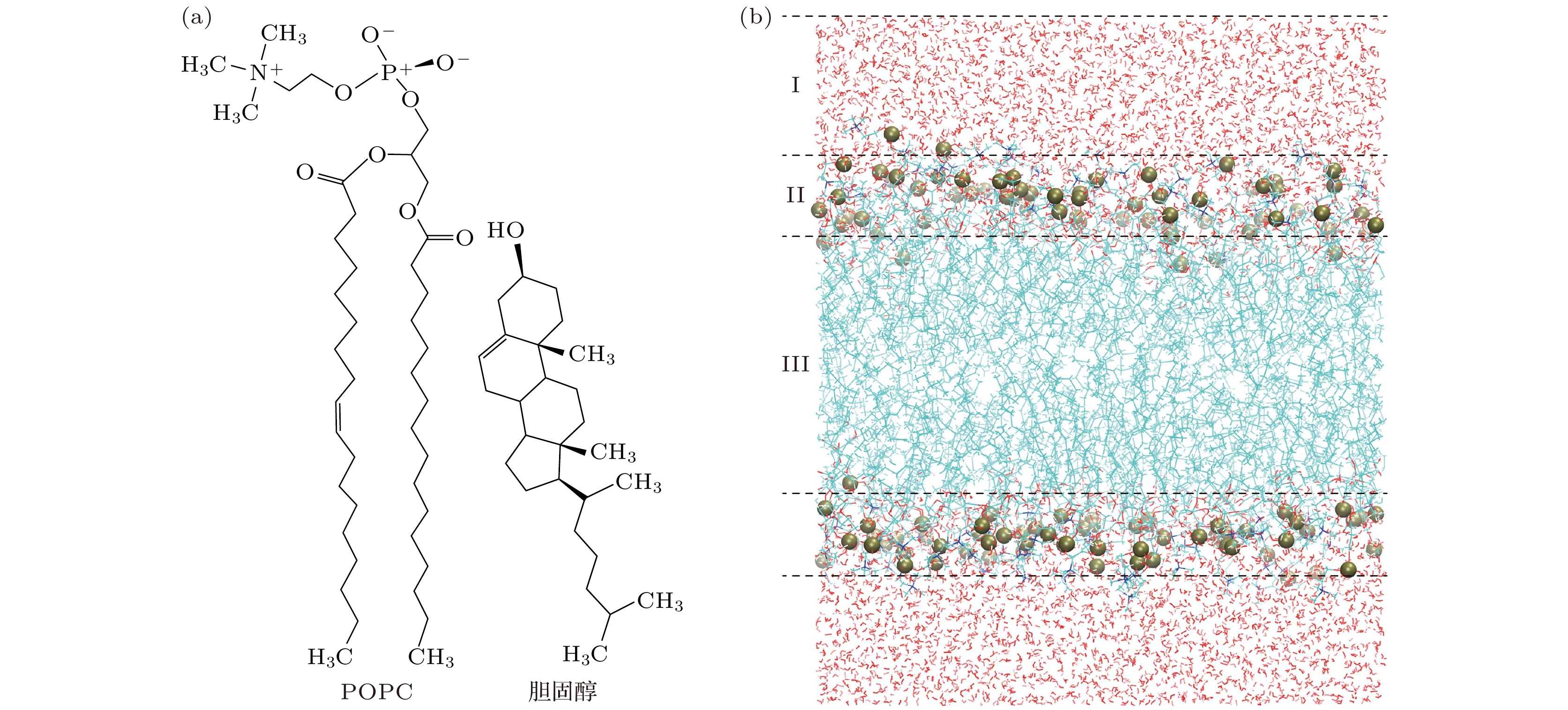
图 1 (a) POPC和胆固醇分子结构; (b) 细胞膜模型示意图. I为液体相, II为POPC头部基团区域, III为POPC尾部区域
Figure 1. (a) Structural components of POPC and cholesterol; (b) schematic diagram of the cell membrane model. I, II, and III represent the liquid phase, the region of POPC head groups and tail groups, respectively.
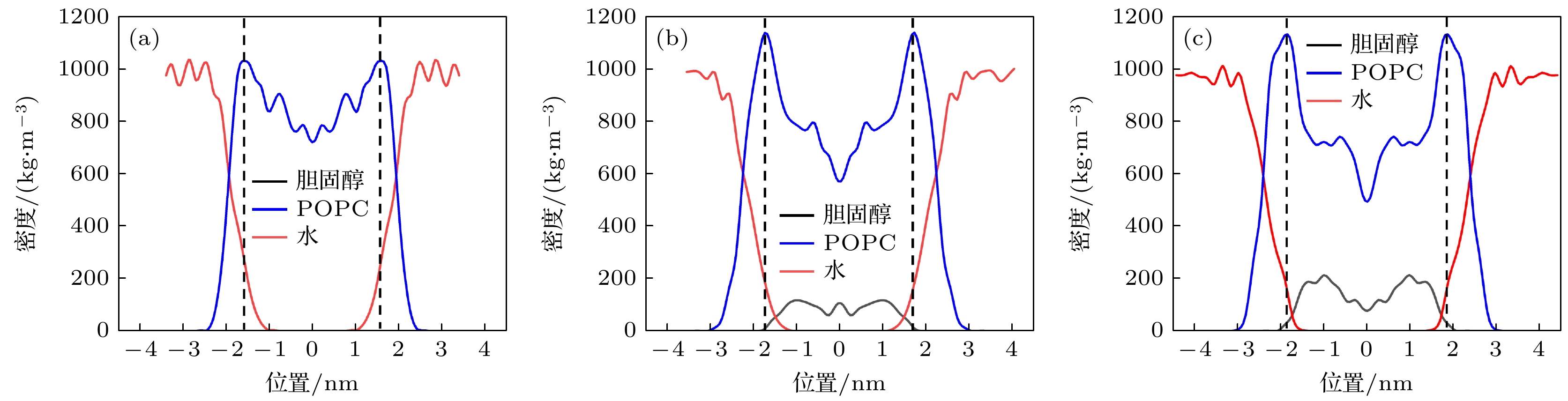
图 2 不同模型的细胞膜密度分布图, 黑色虚线表示两个POPC层的位置 (a)模型1; (b)模型2; (c)模型3
Figure 2. Density distribution of different cell membrane models: (a) Model 1; (b) Model 2; (c) Model 3. The black dashed lines demarcate the interfacial boundaries of the two POPC molecular layers.
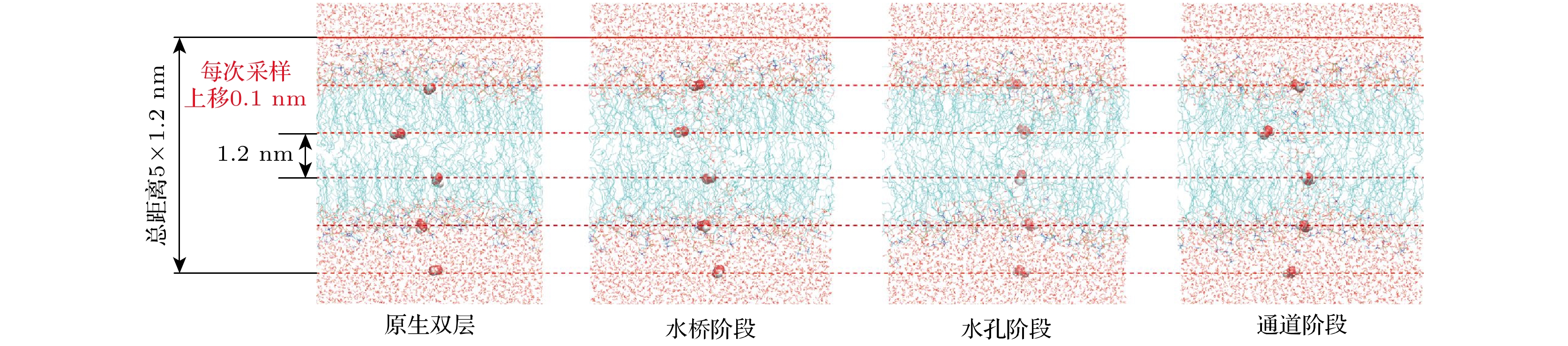
图 3 伞状采样法示意图. 红色虚线表示采样窗口所处的位置
Figure 3. Schematic diagram of umbrella sampling method. The dotted red line indicates the location of the sampling window.
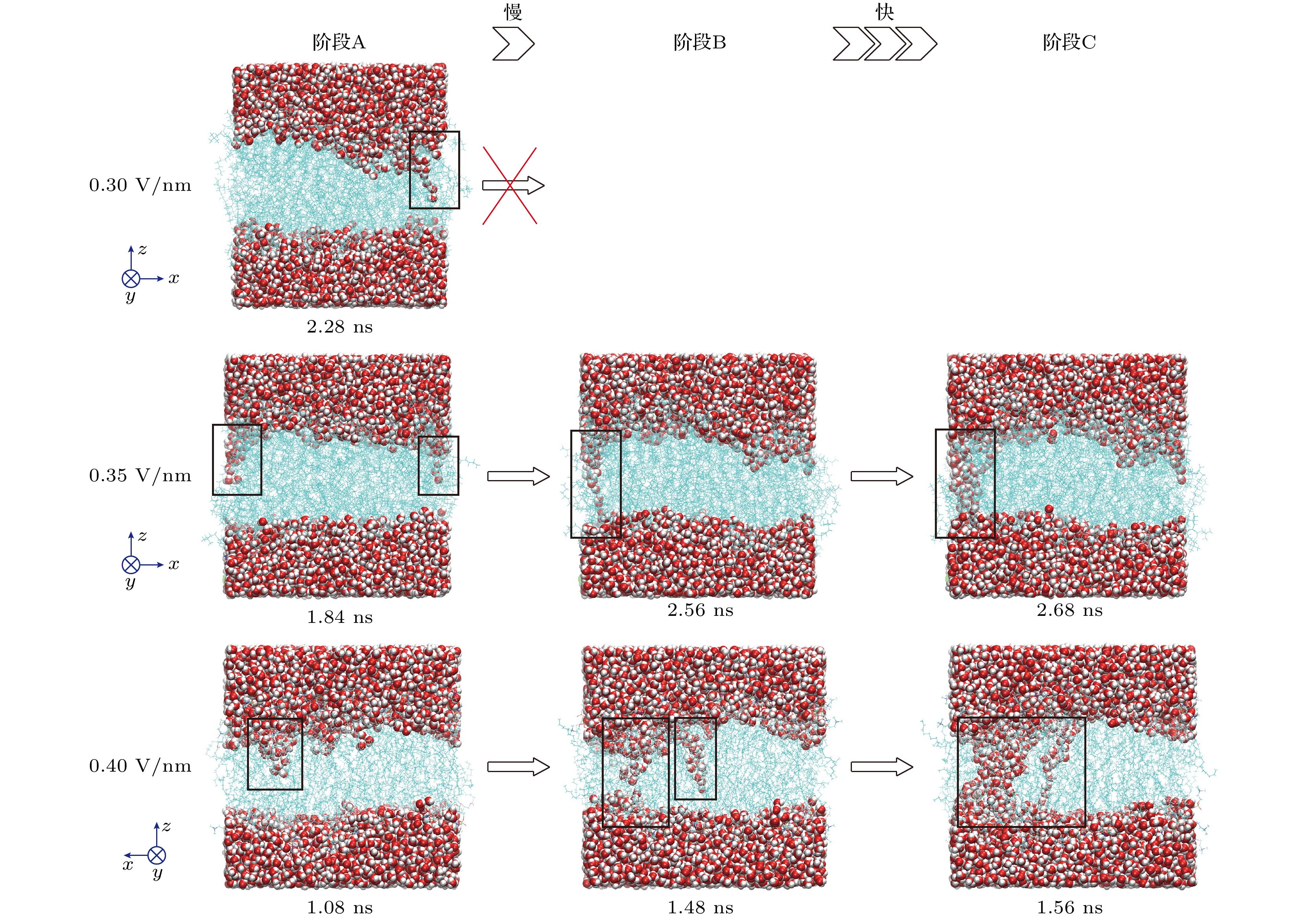
图 4 不同电场强度下细胞膜EP过程示意图. 黑色方框指示出了水突起、水桥和水通道的位置
Figure 4. Schematic diagram of EP processes in cell membrane under different electric field intensities. The black box indicates the location of the water bump, water bridge, and water channel.
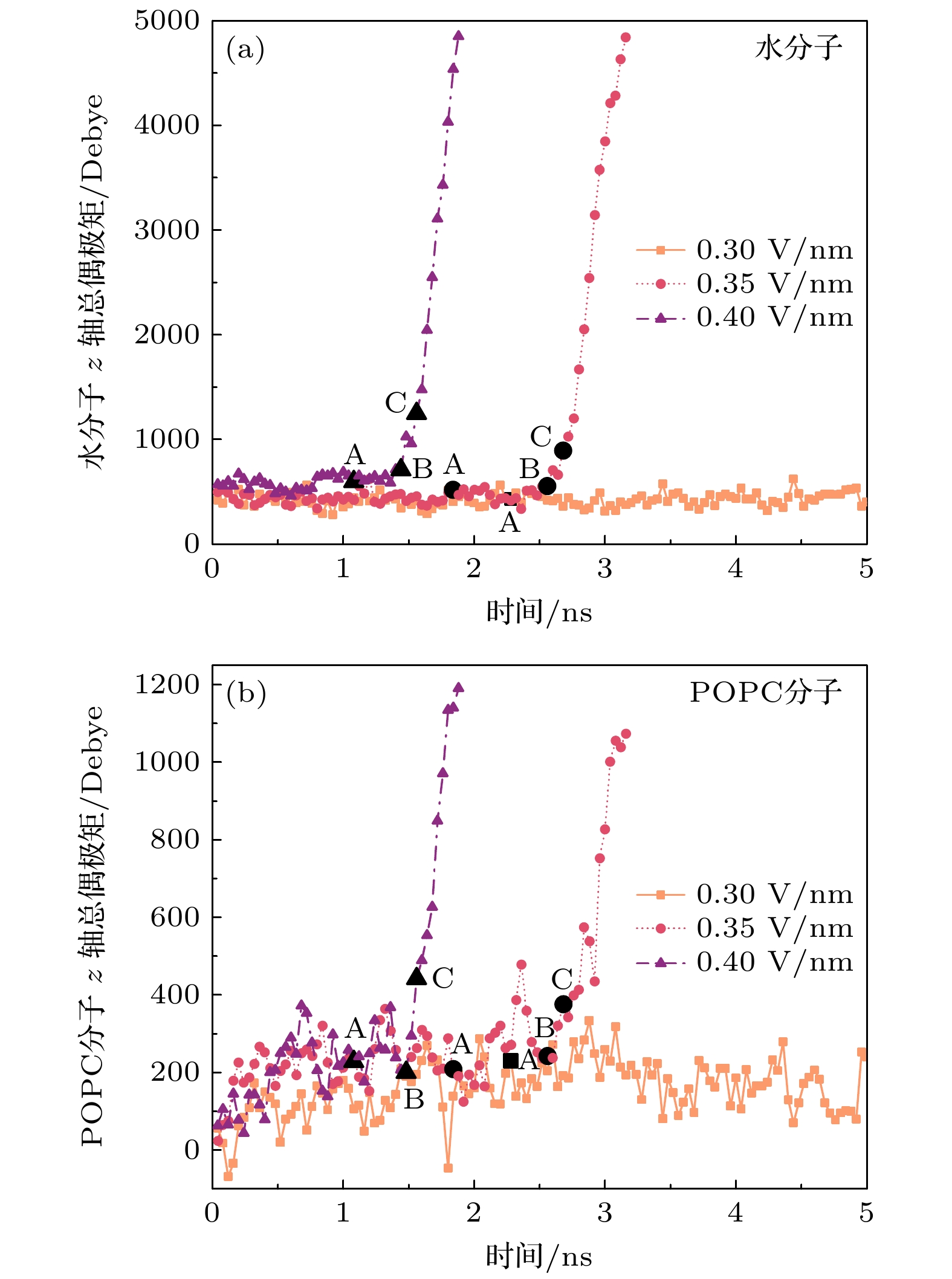
图 5 EP进展过程中细胞膜 (a)水分子和(b) POPC分子的z轴总偶极矩. A, B, C分别表示图4中EP的三个阶段, 1 debye =3.335×10–30 C·m
Figure 5. Mean z-axis dipole moment of (a) water molecules and (b) POPC molecules during EP progression. A, B, and C represents the three stages of EP in Figure 4, respectively. 1 debye =3.335×10–30 C·m.
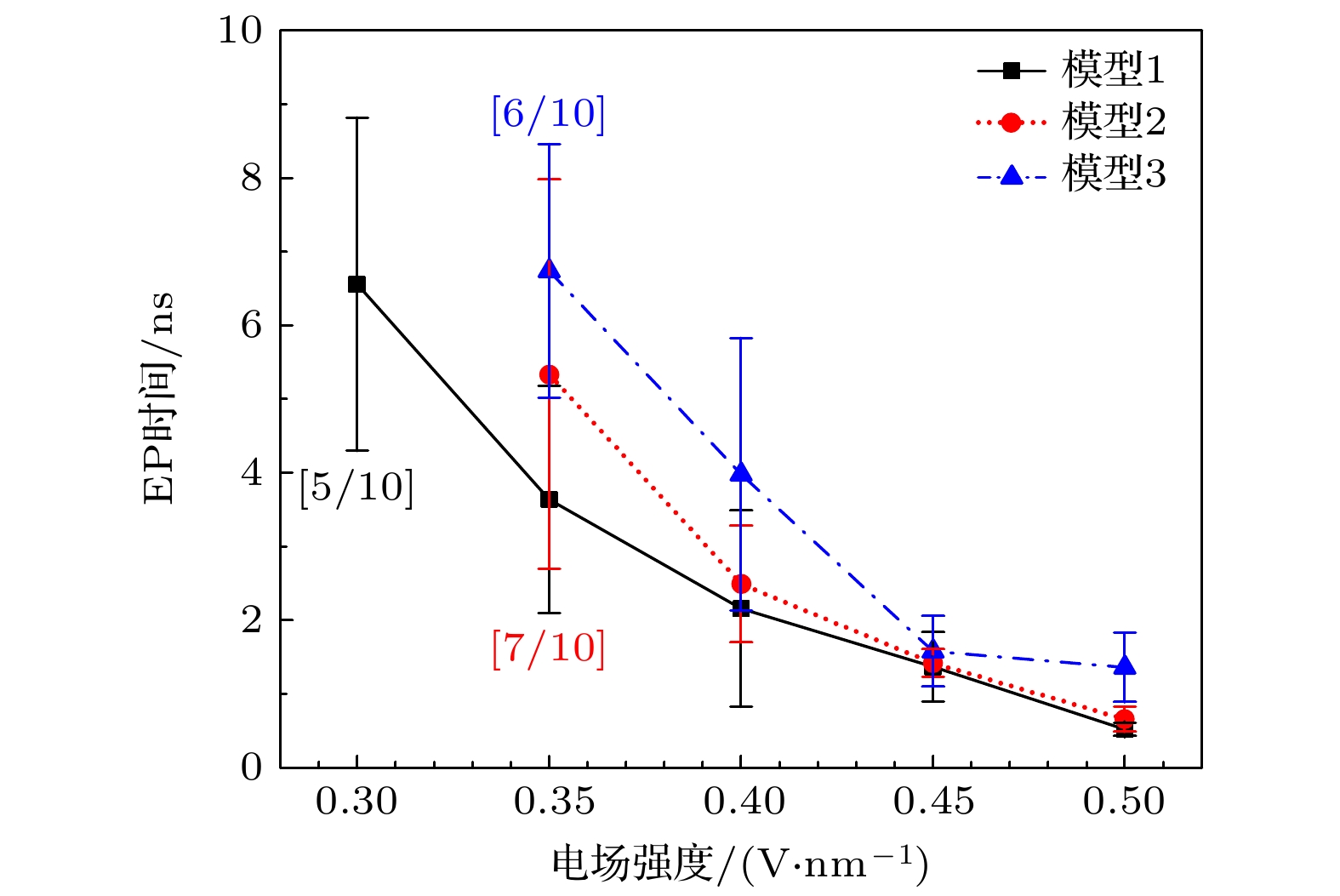
图 6 细胞膜形成EP的时间与电场强度的关系. 括号内分数为: (模拟时间内发生EP的次数)/(总模拟次数)
Figure 6. Relationship between EP formation time and electric field intensity in cell membrane. The score inside the brackets is: (Number of EP occurrences in the simulation time)/(total number of simulations).
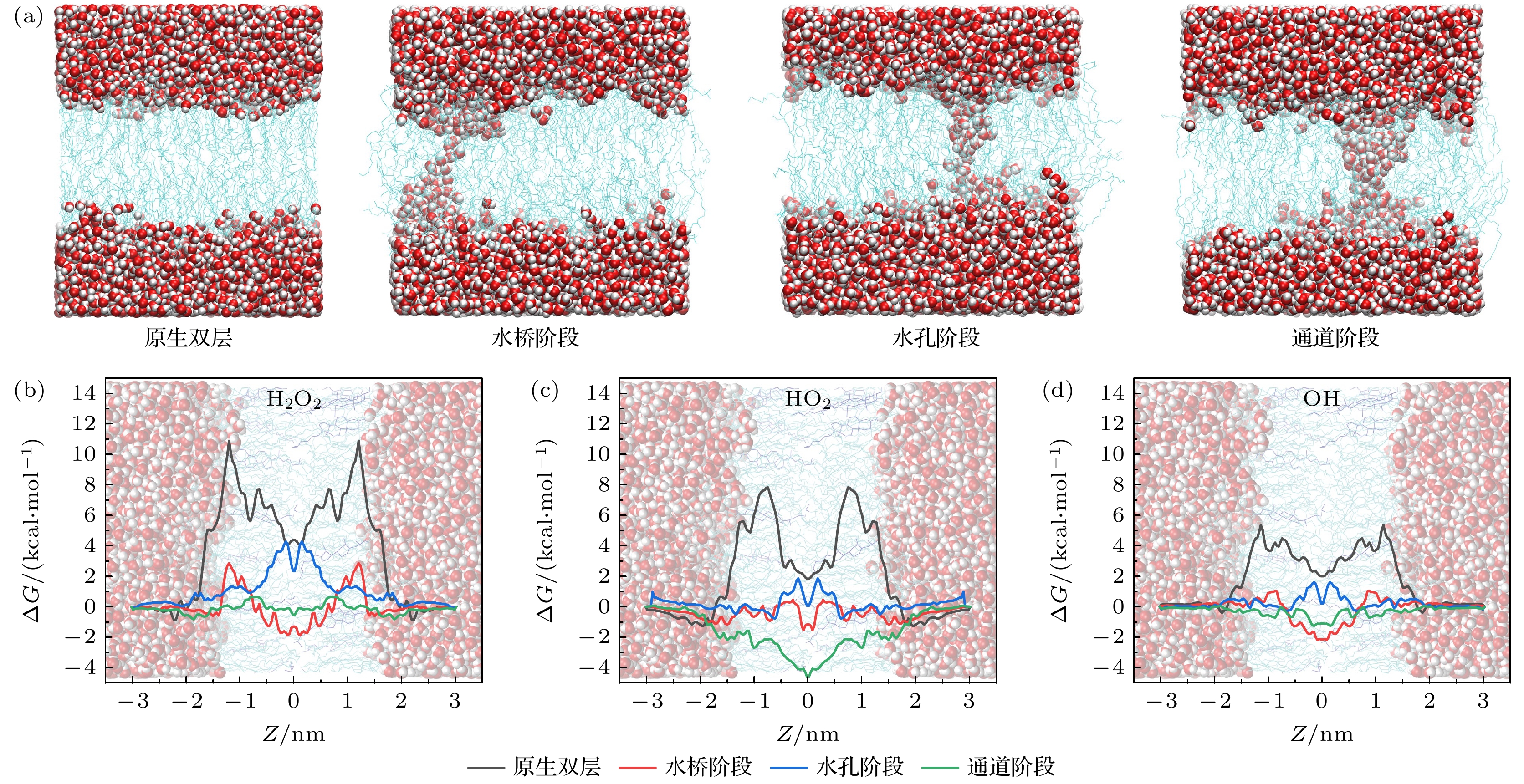
图 7 模型1中ROS在EP不同阶段转运的FEP (a)原生细胞膜构象及EP形成过程中的三个阶段构象; (b) H2O2, (c) HO2和(d) OH的FEP
Figure 7. FEP of ROS transport at different stages of EP in Model 1: (a) The conformation of the native cell membrane and the three-stage conformations during the formation of EP; FEP of (b) H2O2, (c) HO2, and (d) OH.
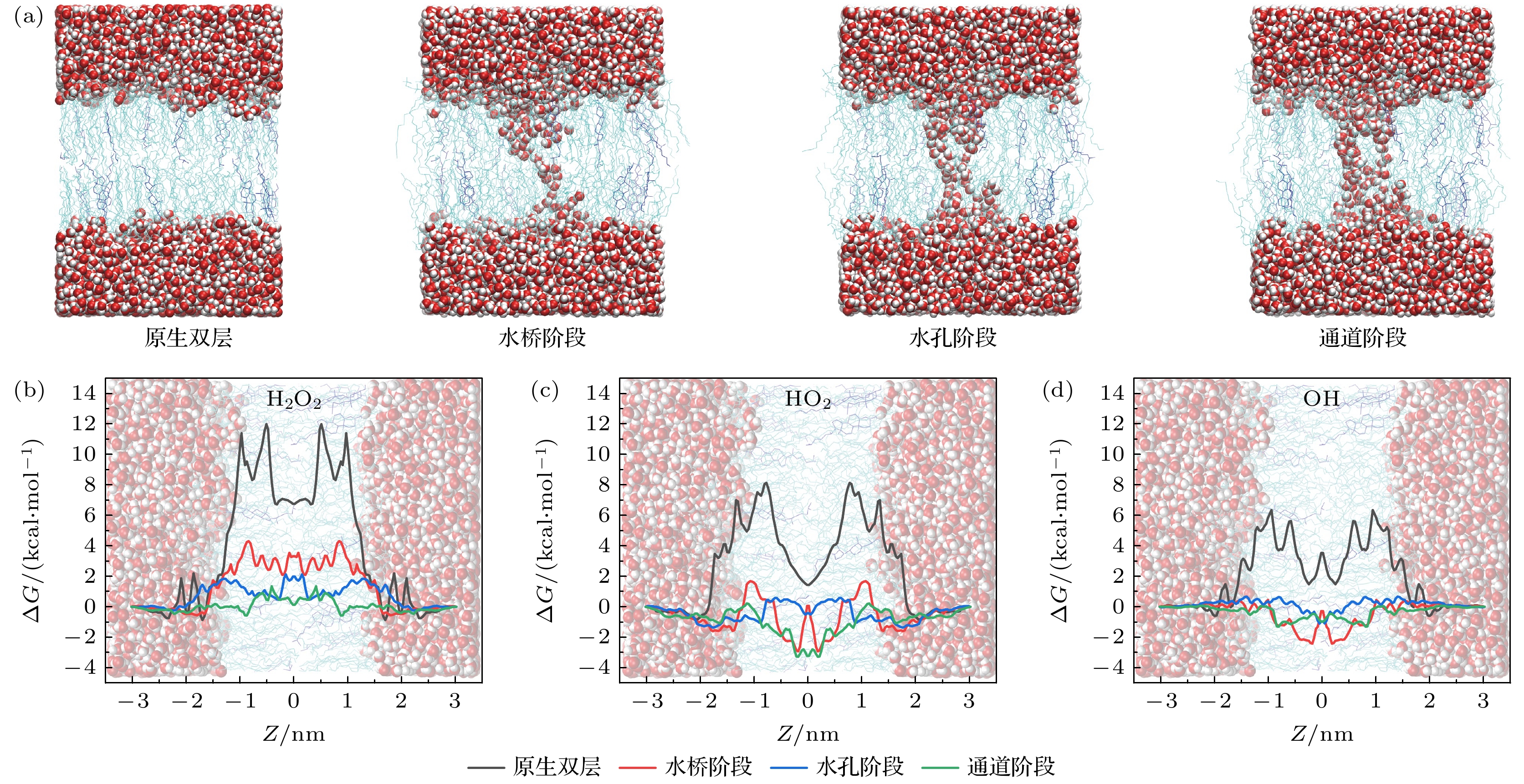
图 8 模型2中ROS在EP不同阶段转运的FEP (a)原生细胞膜构象及EP形成过程中的三个阶段构象; (b) H2O2, (c) HO2和(d) OH的FEP
Figure 8. FEP of ROS transport at different stages of EP in Model 2: (a) The conformation of the native cell membrane and the three-stage conformations during the formation of EP; FEP of (b) H2O2, (c) HO2, and (d) OH.
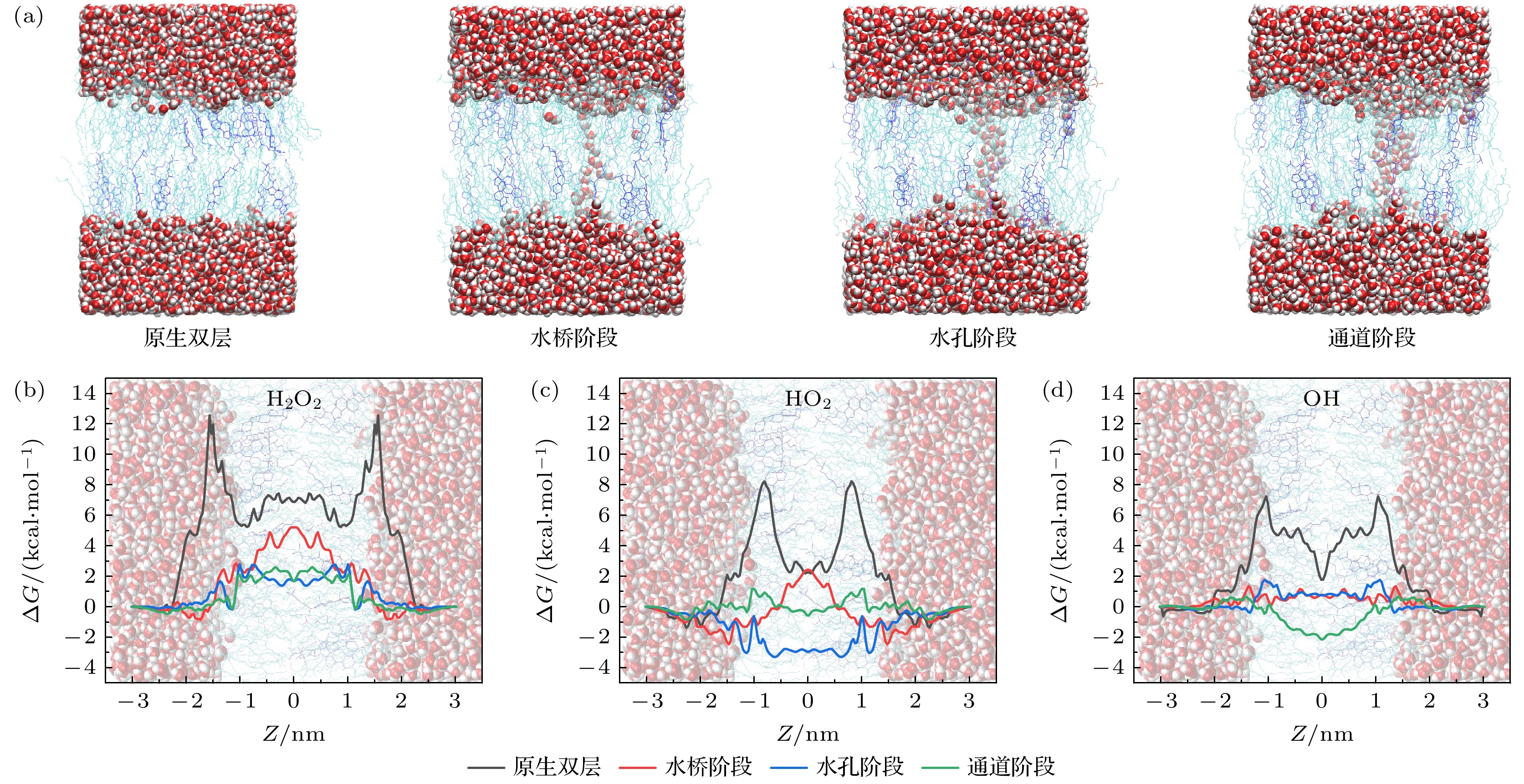
图 9 模型3中ROS在EP不同阶段转运的FEP (a) 原生细胞膜构象及EP形成过程中的三个阶段构象; (b) H2O2, (c) HO2和(d) OH的FEP
Figure 9. FEP of ROS transport at different stages of EP in Model 3: (a) The conformation of the native cell membrane and the three-stage conformations during the formation of EP; FEP of (b) H2O2, (c) HO2, and (d) OH.
表 1 不同胆固醇含量的细胞膜模型参数
Table 1. Cell membrane model parameters with different cholesterol content.
序号 POPC数量 胆固醇数量 胆固醇含量/% 水分子数 X/nm Y/nm Z/nm 模型1 128 0 0 4873 6.03 6.03 9.07 模型2 112 16 12.5 4705 5.94 5.94 8.25 模型3 102 26 20.3 4513 5.78 5.78 8.39  下载: 导出CSV
下载: 导出CSV
-
[1] 卢新培, 罗婧怡, 聂兰兰, 刘大伟, 张冠军, 刘定新, 邵涛, 方志, 金珊珊, 赵亚军, 张远涛, 邹亮, 王晓龙, 李和平, 张宇, 刘东平, 杨德正, 陈支通, 黄青, 程诚, 吴淑群, 刘巧珏, 裴学凯, 闫旭, 程鹤, 熊青, 石琦, 宋珂, 曹颖光, 陈宏翔, 冯爱平, 夏育民, 白帆, 杨春俊, 杨润功, 何光源 2024 高电压技术 50 3555 Lu X P, Luo J Y, Nie L L, Liu D W, Zhang G J, Liu D X, Shao T, Fang Z, Jin S S, Zhao Y J, Zhang Y T, Zou L, Wang X L, Li H P, Zhang Y, Liu D P, Yang D Z, Chen Z T, Huang Q, Chen C, Wu S Q, Liu Q J, Pei X K, Yan X, Cheng H, Xiong Q, Shi Q, Song K, Cao Y G, Chen H X, Feng A P, Xia Y M, Bai F, Yang C J, Yang R G, He G Y 2024 High Voltage Eng. 50 3555 [2] Chen X, Wang X Q, Zhang B X, Yuan M, Yang S Z 2023 Chin. Phys. B 32 115201 doi: 10.1088/1674-1056/ace768 [3] Fang J L, Zhang Y R, Lu C Z, Gu L L, Xu S F, Guo Y, Shi J J 2024 Chin. Phys. B 33 015201 doi: 10.1088/1674-1056/ad0118 [4] Xu H M, Gao J G, Jia P Y, Ran J X, Chen J Y, Li J M 2024 Chin. Phys. B 33 015205 doi: 10.1088/1674-1056/ad0149 [5] Schleusser S, Schulz L, Song J, Deichmann H, Griesmann A C, Stang F H, Mailaender P, Kraemer R, Kleemann M, Kisch T 2022 Microcirculation 29 e12754 doi: 10.1111/micc.12754 [6] Filipic A, Gutierrez-Aguirre I, Primc G, Mozetic M, Dobnik D 2020 Trends Biotechnol. 38 1278 doi: 10.1016/j.tibtech.2020.04.003 [7] Nguyen L, Lu P, Boehm D, Bourke P, Gilmore B F, Hickok N J, Freeman T A 2019 Biol. Chem. 400 77 doi: 10.1515/hsz-2018-0235 [8] Zhou R W, Zhang X H, Zong Z C, Li J X, Yang Z B, Liu D P, Yang S Z 2015 Chin. Phys. B 24 085201 doi: 10.1088/1674-1056/24/8/085201 [9] Borges A C, Kostov K G, Pessoa R S, de Abreu G M A, Lima G d M G, Figueira L W, Koga-Ito C Y 2021 Appl. Sci. 11 1975 doi: 10.3390/app11051975 [10] von Woedtke T, Laroussi M, Gherardi M 2022 Plasma Sources Sci. Technol. 31 054002 doi: 10.1088/1361-6595/ac604f [11] Min T, Xie X, Ren K, Sun T, Wang H, Dang C, Zhang H 2022 Front. Med. 9 884887 doi: 10.3389/fmed.2022.884887 [12] Yan D, Horkowitz A, Wang Q, Keidar M 2021 Plasma Processes Polym. 18 e2100020 doi: 10.1002/ppap.202100020 [13] Yan D, Sherman J H, Keidar M 2017 Oncotarget 8 15977 doi: 10.18632/oncotarget.13304 [14] 姚陈果 2018 高电压技术 44 248 Yao C G 2018 High Voltage Eng. 44 248 [15] Graves D B 2012 J. Phys. D: Appl. Phys. 45 263001 doi: 10.1088/0022-3727/45/26/263001 [16] Haberl S, Miklavcic D, Sersa G, Frey W, Rubinsky B 2013 IEEE Electr. Insul. Mag. 29 29 doi: 10.1109/MEI.2013.6410537 [17] Ruzgys P, Novickij V, Novickij J, Satkauskas S 2019 Bioelectrochemistry 127 87 doi: 10.1016/j.bioelechem.2019.02.002 [18] Wu E, Nie L, Liu D, Lu X, Ostrikov K 2023 Free Radical Biol. Med. 198 109 doi: 10.1016/j.freeradbiomed.2023.02.011 [19] Szlasa W, Kielbik A, Szewczyk A, Rembialkowska N, Novickij V, Tarek M, Saczko J, Kulbacka J 2021 Molecules 26 154 doi: 10.3390/molecules26010154 [20] 孙远昆, 郭良浩, 王凯程, 王少萌, 宫玉彬 2021 物理学报 70 248701 doi: 10.7498/aps.70.20211725 Sun Y K, Guo L H, Wang K C, Wang S M, Gong Y B 2021 Acta Phys. Sin. 70 248701 doi: 10.7498/aps.70.20211725 [21] 邢人芳, 陈明, 李芮羽, 李淑倩, 张瑞, 胡笑钏 2024 物理学报 73 188703 doi: 10.7498/aps.73.20240927 Xing R F, Chen M, Li R Y, Li S Q, Zhang R, Hu X C 2024 Acta Phys. Sin. 73 188703 doi: 10.7498/aps.73.20240927 [22] Hu X, Jin X, Xing R, Liu Y, Feng Y, Lyu Y, Zhang R 2023 Results Phys. 51 106621 doi: 10.1016/j.rinp.2023.106621 [23] Yang S, Zhao T, Zou L, Wang X, Zhang Y 2019 Phys. Plasmas 26 083504 doi: 10.1063/1.5097243 [24] Zhao X, Ding W, Wang H, Wang Y, Liu Y, Li Y, Liu C 2023 J. Chem. Phys. 159 045101 doi: 10.1063/5.0143648 [25] Bera I, Payghan P V 2019 Curr. Pharm. Des. 25 3339 doi: 10.2174/1381612825666190903153043 [26] Arbeitman C R, Rojas P, Ojeda-May P, Garcia M E 2021 Nat. Commun. 12 5407 doi: 10.1038/s41467-021-25478-7 [27] Semmler M L, Bekeschus S, Schäfer M, Bernhardt T, Fischer T, Witzke K, Seebauer C, Rebl H, Grambow E, Vollmar B, Nebe J B, Metelmann H-R, Woedtke T v, Emmert S, Boeckmann L 2020 Cancers 12 269 doi: 10.3390/cancers12020269 [28] Van der Paal J, Neyts E C, Verlackt C C W, Bogaerts A 2016 Chem. Sci. 7 489 doi: 10.1039/C5SC02311D [29] Guo F, Zhou J, Wang J, Qian K, Qu H 2023 Phys. Chem. Chem. Phys. 25 14096 doi: 10.1039/D2CP04637G [30] Bussi G, Donadio D, Parrinello M 2007 J. Chem. Phys. 126 014101 doi: 10.1063/1.2408420 [31] Parrinello M, Rahman A 1981 J. Appl. Phys. 52 7182 doi: 10.1063/1.328693 [32] Hoover W G 1985 Phys. Rev. A 31 1695 doi: 10.1103/PhysRevA.31.1695 [33] Nose S 1984 Mol. Phys. 52 255 doi: 10.1080/00268978400101201 [34] Hess B, Bekker H, Berendsen H J C, Fraaije J 1997 J. Comput. Chem. 18 1463 doi: 10.1002/(SICI)1096-987X(199709)18:12<1463::AID-JCC4>3.0.CO;2-H [35] Darden T, York D, Pedersen L 1993 J. Chem. Phys. 98 10089 doi: 10.1063/1.464397 [36] Yusupov M, Van der Paal J, Neyts E C, Bogaerts A 2017 BBA-Gen. Subjects 1861 839 doi: 10.1016/j.bbagen.2017.01.030 [37] Hu Q, Joshi R P, Schoenbach K H 2005 Phys. Rev. E 72 031902 doi: 10.1103/PhysRevE.72.031902 [38] Hu Q, Viswanadham S, Joshi R P, Schoenbach K H, Beebe S J, Blackmore P F 2005 Phys. Rev. E 71 031914 doi: 10.1103/PhysRevE.71.031914 [39] Schmid N, Eichenberger A P, Choutko A, Riniker S, Winger M, Mark A E, van Gunsteren W F 2011 Eur. Biophys. J. Biophys. 40 843 doi: 10.1007/s00249-011-0700-9 [40] Cordeiro R M, Yusupov M, Razzokov J, Bogaerts A 2020 J. Phys. Chem. B 124 1082 doi: 10.1021/acs.jpcb.9b08172 [41] Neto A J P, Cordeiro R M 2016 BBA-Biomembranes 1858 2191 doi: 10.1016/j.bbamem.2016.06.018 [42] Razzokov J, Yusupov M, Cordeiro R M, Bogaerts A 2018 J. Phys. D: Appl. Phys. 51 365203 doi: 10.1088/1361-6463/aad524 [43] Wu S Q, Dong X, Pei X K, Yue Y F, Lu X P 2017 Trans. Chin. Electrotech. Soc. 32 82 (in Chinse) [吴淑群, 董熙, 裴学凯, 岳远富, 卢新培 2017 电工技术学报 32 82] Wu S Q, Dong X, Pei X K, Yue Y F, Lu X P 2017 Trans. Chin. Electrotech. Soc. 32 82 (in Chinse) [44] Nakagawa Y, Ono R, Oda T 2011 J. Appl. Phys. 110 073304 doi: 10.1063/1.3638457 [45] Verreycken T, van der Horst R M, Baede A H F M, Van Veldhuizen E M, Bruggeman P J 2012 J. Phys. D: Appl. Phys. 45 045205 doi: 10.1088/0022-3727/45/4/045205 [46] Vermeylen S, Waele J D, Vanuytsel S, Backer J D, Van de Paal J, Ramakers M, Leyssens K, Marcq E, Van Audenaerde J, Smiths E L J, Dewilde S, Bogaerts A 2016 Plasma Processes Polym. 13 1195 doi: 10.1002/ppap.201600116 [47] Kim S J, Seong M J, Mun J J, Bae J H, Joh H M, Chung T H 2022 Int. J. Mol. Sci. 23 14092 doi: 10.3390/ijms232214092 [48] Geboers B, Scheffer H J, Graybill P M, Ruarus A H, Nieuwenhuizen S, Puijk R S, van den Tol P M, Davalos R V, Rubinsky B, de Gruijl T D, Miklavcic D, Meijerink M R 2020 Radiology 295 254 doi: 10.1148/radiol.2020192190 [49] Jiang C L, Davalos R V, Bischof J C 2015 IEEE Trans. Biomed. Eng. 62 4 doi: 10.1109/TBME.2014.2367543 -
计量
- 文章访问数: 194
- HTML全文浏览数: 194
- PDF下载数: 4
- 施引文献: 0