
 首页
首页 登录
登录 注册
注册



-
炎症是对感染、创伤或组织损伤的急性反应,清除致病性物质从而保护机体,炎症可能出现在心脑血管、糖尿病和骨质疏松症等疾病的任意时期[1]。在炎症过程中,许多关键的转录因子被激活,如NF-κβ信号通路进一步诱导IL-6、TNF-α等炎症因子释放;转录因子对转运体和代谢酶的调节起着关键作用[2-4],导致体内代谢紊乱,影响药物的血药浓度和治疗作用。因此,研究炎症状态下体内的代谢改变具有重要意义。
脂多糖(LPS)是革兰氏阴性菌外膜的组成部分,进入血液后会导致全身的炎症[5],与临床炎症相似,是经典的炎症造模药物,常被用来建立动物的炎症模型或诱导细胞炎症[6-7]。Puris等[8]研究了LPS诱导小鼠全身炎症对小鼠血浆、脑皮质和海马靶向代谢组、脂质组以及参与的脑皮质蛋白表达的短期影响,发现皮质和海马中的磷脂酰胆碱和磷脂酰乙醇胺水平显著增加,脑皮质中甜菜碱水平显著降低,小鼠大脑皮层 GluN1 受体蛋白表达显著增加。
新西兰白兔对LPS敏感,本研究拟利用实验室前期建立的兔7天炎症急性反应模型方法[9],在造模的第2天及第7天收集兔尿液,采用超高效液相色谱-高分辨质谱(UPLC-HRMS)法进行炎症急性反应期代谢组学研究,分析差异代谢通路,以寻找急性炎症反应期的作用靶点,解释其对药代动力学的影响机制,希望为炎症期间疾病的治疗提供信息。
-
AB SCIEX ZenoTOFTM 7600高分辨质谱仪:美国AB Sciex公司产品;Acquity UPLC液相色谱系统:美国Waters公司产品;Centrifuge 5425 高速冷冻离心机:德国Eppendorf公司产品。
-
乙腈(色谱纯):德国Merck公司产品;甲酸(色谱纯):德国Sigma公司产品;超纯水:杭州娃哈哈集团有限公司产品;脂多糖:美国Sigma公司产品,以生理盐水作为溶剂。
-
8只新西兰兔:购自绿叶制药有限公司实验动物中心,体重3~4 kg,常规饲养于SPF级动物房,自由进食进水。炎症急性反应期模型采用耳缘静脉注射递增剂量LPS,第1~7天给予LPS剂量分别为0.1、0.2、0.5、1、1.5、2、2 μg/kg,给药体积为0.5 mL/kg。在给予LPS前、给予LPS 2天、给予 LPS 7天进行12 h尿液收集(早晨8:00~晚上8:00),分别作为空白组(B)、造模2天组(T)和造模7天组(S)。将尿液样本在4 ℃以3 000 r/min离心10 min,取上清液于Eppendorf(EP)管内,−80 ℃冰箱冻存,待后续分析。检测前,在4 ℃冰箱解冻尿液样品,于4 ℃以13 500 r/min离心 10 min,取上清液进行检测。所有样品取等体积上清液制成6个质控(QC)样品,在相同条件下穿插在样品分析过程中检测。
-
Kinetex F5色谱柱(150 mm×2.1 mm×2.6 μm);流动相:A为0.05%甲酸-水溶液,B为0.05%甲酸-乙腈溶液;梯度洗脱程序:0~1.5 min(0%~2%B),1.5~3 min(2%~25%B),3~13 min(25%~60%B),13~15 min(60%~95%B),15~17 min(95%B),17~17.1 min(95%~2%B);流速0.3 mL/min,柱温35 ℃,进样体积1 μL。
-
采用正、负离子扫描模式,雾化气压强 379 kPa,辅助气压强379 kPa,气帘气压强206 kPa,离子源温度550 ℃,喷雾电压5 000 V(正离子模式)/−4 500 V(负离子模式),去簇电压60 V,碰撞能量(35±15) eV,质量扫描范围m/z 60~1 000。通过信息依赖采集(IDA)和动态背景扣除(DBS)功能获得高分辨质谱数据。
-
利用R语言软件包对原始谱图进行峰提取,SCIEX OS软件的本地数据库和MetDNA网络数据库对二级质谱数据进行匹配。使用MarkerView™软件进行主成分分析(PCA)和偏最小二乘法判别分析(PLS-DA),得到变量重要性投影(VIP)值、显著性(p)和差异倍数(FC)值。利用metaboAnalyst 5.0(https://www.metaboanalyst.ca/)进行代谢通路分析,使用KEGG数据库筛选出具有显著差异的代谢通路。
-
在分析批数据采集过程中穿插QC样品测定,正、负离子模式下,QC样品的保留时间和信号强度的重现性良好,示于图1a、1b。对待测样本及QC样品进行PCA 分析,结果示于图1c、1d,可以看出,QC均聚集在一起,表明数据稳定性和重现性较好。
-
排除QC并对B、S、T组样本的数据进行PCA分析,结果表明,在正、负离子模式下,各组间分类趋势不明显,示于图2a、2b,其原因是生物样本组内、组间存在变异,仅通过PCA难以得到显著的组间分类效果。为了使组间差异更显著,使用 PLS-DA进行分析,结果示于图2c、2d。可见,造模2天组和造模7天组与空白组对比均分类明显。PLS-DA对造模组和空白组的区分能力优于PCA,有助于发现LPS诱导兔炎症急性模型体内的代谢特性变化。
-
对空白组和造模组进行差异代谢物分析,差异代谢物筛选标准为同时满足VIP>1、p<0.05和FC>2。与空白组相比,造模 2天组尿液中共发现41个差异代谢物,其中正、负离子模式分别为22、19个;造模7天组共发现161个差异代谢物,其中正、负离子模式分别为53、108个。通过聚类热图分析差异代谢物,结果示于图3,发现造模2天组和造模7天组与空白组之间存在聚类相关性,且代谢物在造模组中高表达。
-
将筛选得到的差异代谢物进行代谢通路分析,具有显著性差异(p<0.01)的代谢通路主要为类固醇激素生物合成(steroid hormone biosynthesis)通路,在造模2天组和7天组,p值均小于0.01;而其他代谢通路差异并不明显,示于图4。与空白组相比,在类固醇激素生物合成通路上,造模2天组和7天组中分别有15、14个代谢物处于上调趋势,结果列于表1。
-
采用低剂量递增注射LPS方法建立新西兰白兔炎症急性反应期模型,在此期间,新西兰白兔的肝肾功能正常[9],能很好地模拟临床炎症急性反应期状态。通过以上研究发现,类固醇激素生物合成为兔炎症期间的主要差异代谢通路。
类固醇激素通过肾上腺皮质、性腺和胎盘生成[10-11]。胆固醇可以生成孕烯醇酮,是类固醇激素生物合成的前体[12]。孕烯醇酮通过抑制破骨细胞的形成来保护小鼠免受LPS诱导的体内炎症性骨破坏[13]。硫酸脱氢表雄酮和孕烯醇酮可以抑制LPS诱导的肿瘤坏死因子(TNF)的产生[14]。皮质醇具有抗炎作用[15]。实验表明,给药LPS后,尿液中硫酸脱氢表雄酮、孕烯醇酮和皮质醇均出现显著升高,可能是机体自发性调节LPS给药后带来的炎症反应。
类固醇激素代谢主要通过细胞色素P450(CYP450)酶和羟基类固醇脱氢(HSD)酶完成[16-17]。当机体产生炎症时,CYP450和HSD会参与合成类固醇激素以达到抗炎目的,同时,CYP450酶的活性会受到炎症因子的调控[18]。该研究提供了急性炎症期有关靶标的验证,同时在候选药物的测试期间为其提供使用急性炎症模型的重要信息。
-
本研究采用UPLC-HRMS法对LPS诱导的兔炎症急性反应期模型进行尿液非靶向代谢组学研究,在造模 2天组和造模7天组分别发现41和161个差异代谢物,主要为类固醇激素生物合成通路。孕烯醇酮等14个代谢物在类固醇激素生物合成通路处于上调趋势,这为炎症急性反应期有关靶标验证和疾病治疗提供了重要信息,有利于进一步理解炎症反应机制及候选药物评价。
采用高效液相色谱-高分辨质谱研究脂多糖诱导的兔炎症急性期模型代谢组学
Metabolomic Changes of Lipopolysaccharide-Induced Acute-Phase Response Rabbit Inflammation Model Using UPLC-HRMS
-
摘要: 本研究对新西兰白兔低剂量递增给予脂多糖(lipopolysaccharide, LPS),诱导炎症急性反应期模型,收集给予LPS前、给予LPS 2天和7天后的尿液样本,采用超高效液相色谱-高分辨质谱(UPLC-HRMS)技术检测,并进行非靶向代谢组学分析。利用主成分分析(PCA)和偏最小二乘法判别分析(PLS-DA)法筛选差异代谢物,并通过KEGG数据库进行差异代谢通路分析。结果表明,给予LPS 2天和7天后,白兔尿液中的差异代谢物与给药前相比有显著差异,分别鉴定出41和161个差异代谢物,主要与类固醇激素生物合成通路显著相关(p<0.01)。炎症急性反应期发生了代谢紊乱,类固醇激素生物合成通路上调,这为炎症急性反应期有关靶标验证和疾病治疗提供了信息。
-
关键词:
- 代谢组学 /
- 脂多糖(LPS) /
- 差异代谢物 /
- 类固醇激素 /
- 高效液相色谱-高分辨质谱(UPLC-HRMS)
Abstract: Inflammation can occur at any time in many diseases, and affect the activity of metabolism enzymes and transporters, so change the blood concentration and therapeutic effect of therapeutic drugs. Therefore, it is necessary to study the metabolic changes during the acute-phase response (APR) of inflammation. In this study, urine metabolomics was performed to investigate the possible differential metabolic pathways in New Zealand white rabbit APR inflammation model. An APR rabbit model with normal hepatic and renal function was induced by intravenous injection of lipopolysaccharide (LPS) in low-dose escalation, at 0.1, 0.2, 0.5, 1, 1.5, 2 and 2 μg/kg from 1 to 7 days. The urine samples were collected at prior of dosing, 2-day and 7-day after the first dose of LPS, and detected using ultra-high liquid chromatography coupled with high resolution mass spectrometry (UPLC-HRMS). The samples were separated by a Kinetex F5 column (150 mm×2.1 mm×2.6 μm) with the mobile phase of water containing 0.05% formic acid and acetonitrile containing 0.05% formic acid. ZenoTOFTM 7600 HRMS was operated under positive and negative ion modes to collect data, and the high-resolution mass spectral data was acquired by utilizing information dependent acquisition (IDA) and dynamic background subtraction (DBS). Non-targeted metabolomics analysis was completed by principal component analysis (PCA) and partial least squares discriminant analysis (PLS-DA) to search for differential metabolites. Then the differential metabolic pathway analysis was carried out using the metaboAnalyst 5.0 website and KEGG database. The analysis results indicated that the method has high stability and reproducibility with quality control samples clustered together in PCA. The quality control and APR model groups can be separated well by OPLS-DA, which showed that the metabolism in rabbits is significantly disturbed during APR period. Clustering heatmap analysis was performed on the screened differential metabolites which were judged by variable importance projection (VIP>1), significance (p<0.05), and folds of change (FC>2). The results showed differential metabolites in urine at 2-day and 7-day after LPS administration are significantly different from those before administration, 41 and 161 differential metabolites are identified in the two groups of samples, respectively. Furtherly, the differential metabolic pathway analysis revealed that the differential metabolites are significantly correlated with the steroid hormone biosynthesis pathway (p<0.01). Pregnenolone, 11-deoxycorticosterone, 17α,21-dihydroxypregnenolone, 11-deoxycortisol, dehydroepiandrosterone sulphate, 19-oxosteroids, cortisol, 16α-hydroxyandrost-4-ene-3,17-dione, epinephrine, 19-oxoandrost-4-ene-3,17-dione, cortisone, 11β-hydroxyprogesterone, 18-hydroxycorticosterone, 11-dehydrocorticosterone, estrone glucosinolate, and corticosterone are in an up-regulated trend on steroid hormone biosynthesis pathway. The study provides the important information for target verification and disease treatment during the APR of inflammation, and facilitates further understanding of inflammatory response mechanisms. -
-
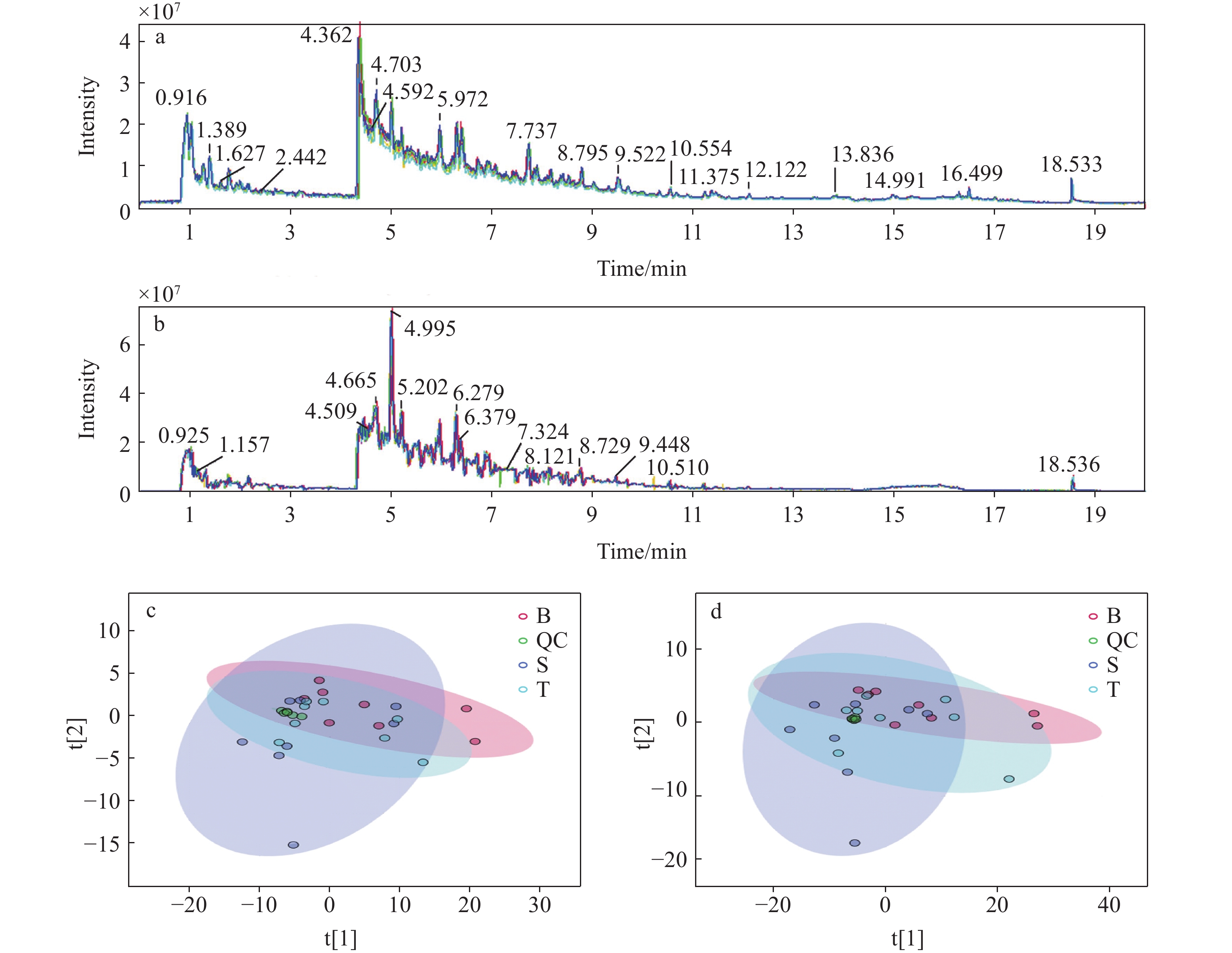
图 1 QC样本的总离子流图(a,b)和QC、空白组、造模2天组和造模7天组PCA图(c,d)
Figure 1. Total ion chromatograms of quality control (QC) sample (a, b), PCA plots of QC, blank, 2-day and 7-day modeling groups (c, d)
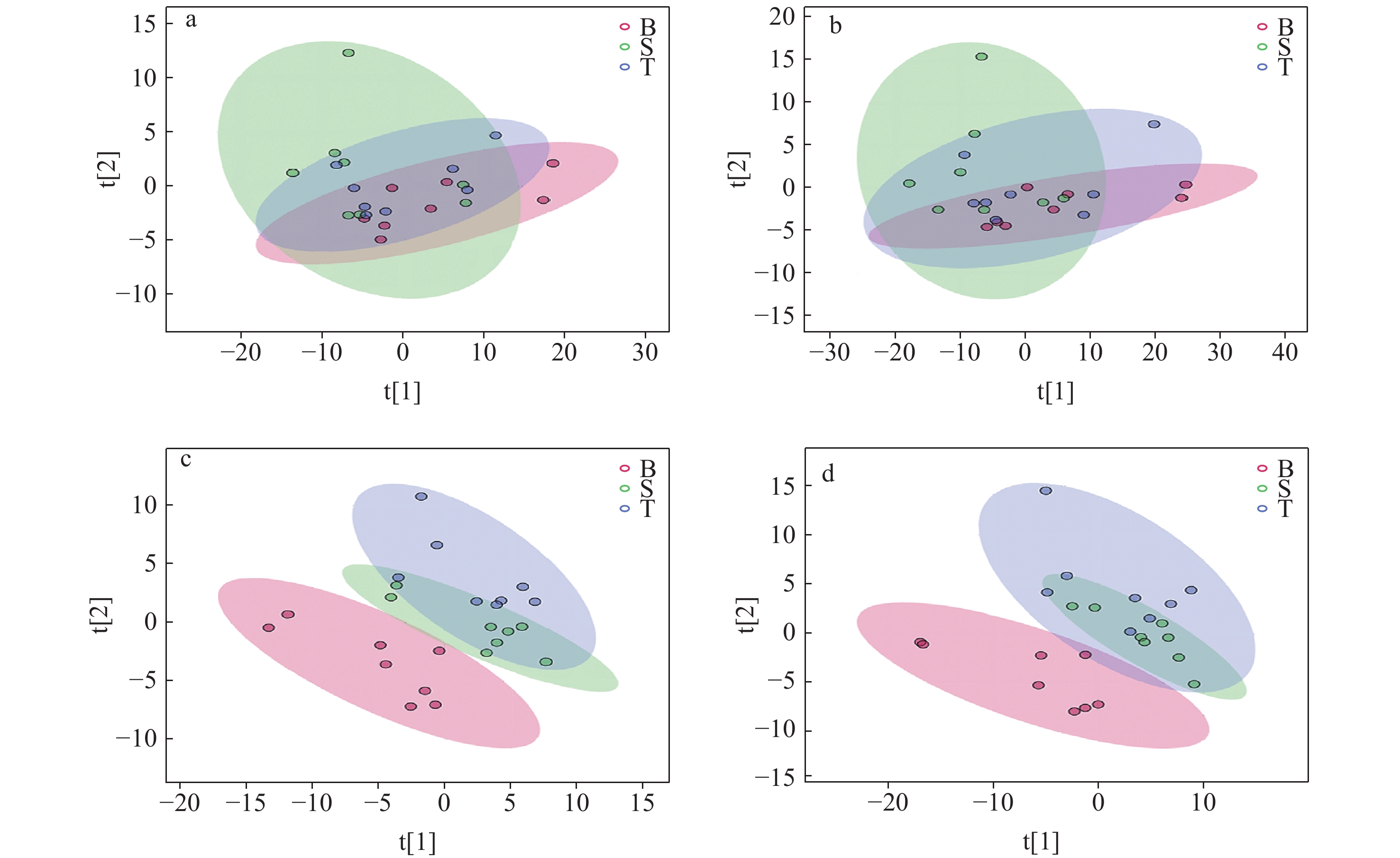
图 2 空白组、造模2天组和造模7天组在正、负离子模式下的PCA图(a, b)和PLS-DA图(c, d)
Figure 2. PCA (a, b) and PLS-DA (c, d) plots of blank, 2-day and 7-day modeling groups under positive and negative ion modes
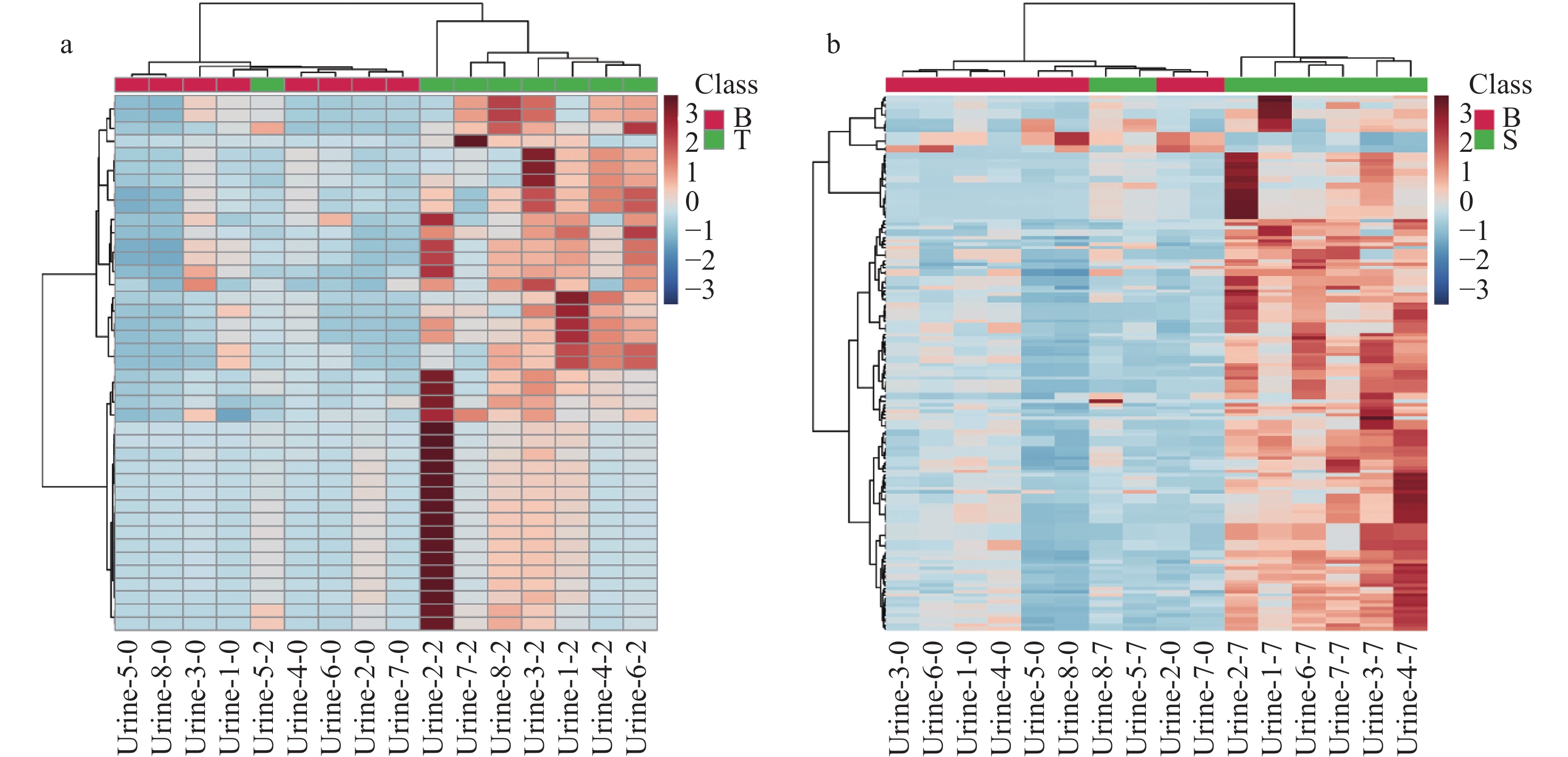
图 3 造模2天组(a)和造模7天组(b)差异代谢物的聚类分析热图
Figure 3. Clustering heat maps of differential metabolites in 2-day (a) and 7-day (b) modeling groups
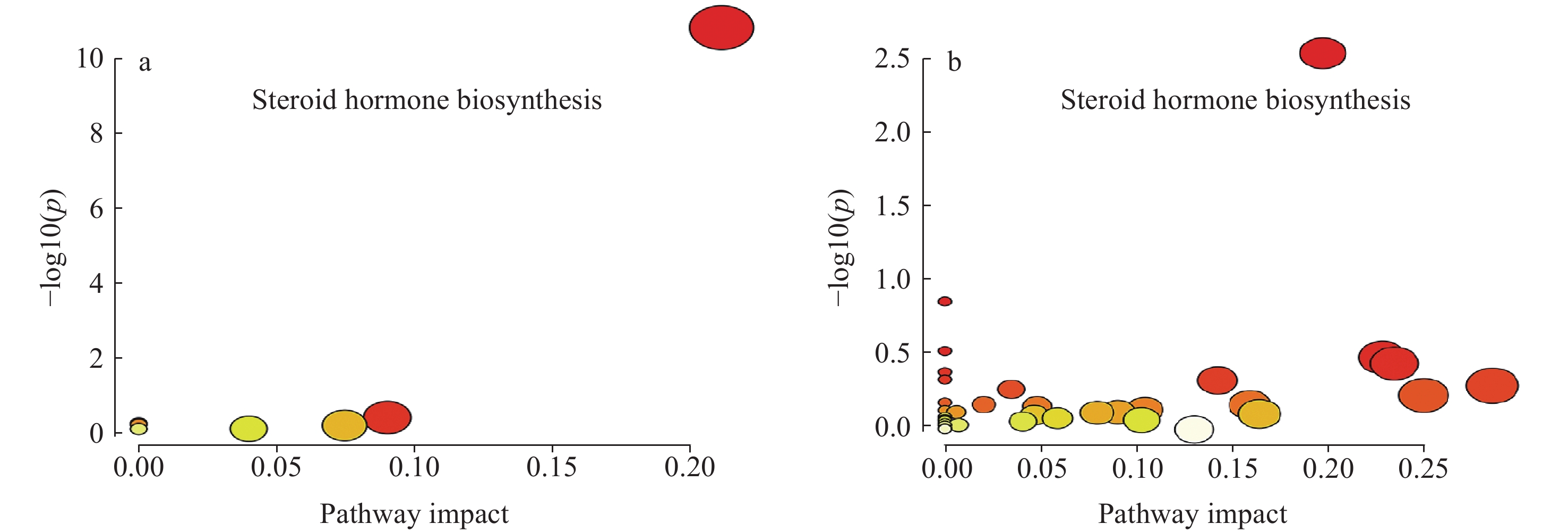
图 4 造模2天组(a)和造模7天组(b)的代谢通路分析
Figure 4. Metabolic pathway analysis of 2-day (a) and 7-day (b) modeling groups
表 1 类固醇激素生物合成通路的差异代谢物
Table 1. Differential metabolites of steroid hormone biosynthesis pathway
分组
Group代谢物
Metabolite分子式
Molecular
formula准分子离子
Quasi-molecular
ion质荷比
Mass charge ratio
(m/z)特征碎片离子
Characteristic fragment ion
(m/z)差异
倍数
FC显著性
p造模
2天孕烯醇酮 C21H32O2 [M+H]+ 317.25 281.22, 159.10 3.63 0.003 11-脱氧皮质酮 C21H30O3 [M+H]+ 331.23 109.06, 97.06 3.60 0.024 17α,21-二羟基孕烯醇酮 C11H32O4 [M+H]+ 349.24 281.22 3.14 0.047 11-脱氧皮质醇 C21H30O4 [M−H]− 345.21 327.21, 121.06 5.09 0.015 硫酸脱氢表雄酮 C19H28O5S [M+CH3COOH−H]− 427.18 97.30 18.6 <0.001 19-氧代甾酮 C19H26O3 [M+CH3COOH−H]− 361.20 117.46 8.26 0.003 皮质醇 C21H30O5 [M+H]+ 363.22 327.20, 121.06 6.38 <0.001 16α-羟基雄甾-4-烯-3,17-二酮 C19H26O3 [M+CH3COOH−H]− 361.20 173.99 8.36 0.003 肾上腺素 C19H24O3 [M+CH3COOH−H]− 359.19 121.20 8.15 0.003 19-氧代雄甾-4-烯-3,17-二酮 C19H24O3 [M+CH3COOH−H]− 359.19 169.60 8.15 0.003 可的松 C21H28O5 [M+H]+ 361.20 163.11, 121.06 8.38 0.002 11β-羟孕酮 C21H30O3 [M+H]+ 331.23 313.22, 121.06 3.60 0.024 18-羟基皮质酮 C21H30O5 [M+H]+ 363.22 191.01 6.38 <0.001 11-脱氢皮质酮 C21H28O4 [M+H]+ 345.21 327.14 7.14 <0.001 雌酮葡糖苷酸 C24H30O8 [M−H2O+H]+ 427.18 269.13 19.1 <0.001 造模
7天孕烯醇酮 C21H32O2 [M+H]+ 317.25 281.22, 159.10 3.31 0.028 17α,21-二羟基孕烯醇酮 C11H32O4 [M+H]+ 349.24 281.22 2.44 0.049 11-脱氧皮质醇 C21H30O4 [M+H]+ 347.22 329.21, 119.06 2.90 0.021 硫酸脱氢表雄酮 C19H28O5S [M+CH3COOH−H]− 427.18 97.30 35.3 <0.001 19-氧代甾酮 C19H26O3 [M+CH3COOH−H]− 361.20 117.46 18.7 <0.001 皮质醇 C21H30O5 [M+H]+ 363.22 327.20, 121.06 15.3 <0.001 16α-羟基雄甾-4-烯-3,17-二酮 C19H26O3 [M+CH3COOH−H]− 361.20 173.99 18.7 <0.001 肾上腺素 C19H24O3 [M+CH3COOH−H]− 359.19 121.20 14.4 <0.001 19-氧代雄甾-4-烯-3,17-二酮 C19H24O3 [M+CH3COOH−H]− 359.19 169.60 14.4 <0.001 可的松 C21H28O5 [M+H]+ 361.20 163.11, 121.06 14.5 <0.001 皮质酮 C21H30O4 [M+H]+ 347.22 109.06, 97.06 2.90 0.021 18-羟基皮质酮 C21H30O5 [M+H]+ 363.22 191.01 15.3 <0.001 11-脱氢皮质酮 C21H28O4 [M+H]+ 345.21 327.14 14.4 <0.001 雌酮葡糖苷酸 C24H30O8 [M−H2O+H]+ 427.18 269.13 36.4 <0.001  下载: 导出CSV
下载: 导出CSV
-
[1] PANIGRAHY D, GILLIGAN M M, SERHAN C N, KASHFI K. Resolution of inflammation: an organizing principle in biology and medicine[J]. Pharmacology & Therapeutics, 2021, 227: 107 879. [2] ERDMANN P, BRUCKMUELLER H, MARTIN P, BUSCH D, HAENISCH S, MÜLLER J, WIECHOWSKA-KOZLOWSKA A, PARTECKE L I, HEIDECKE C D, CASCORBI I, DROZDZIK M, OSWALD S. Dysregulation of mucosal membrane transporters and drug-metabolizing enzymes in ulcerative colitis[J]. Journal of Pharmaceutical Sciences, 2019, 108(2): 1 035-1 046. [3] PAN W, YU C, HSUCHOU H, KASTIN A J. The role of cerebral vascular NFkappaB in LPS-induced inflammation: differential regulation of efflux transporter and transporting cytokine receptors[J]. Cellular Physiology and Biochemistry, 2010, 25(6): 623 -630 . doi: 10.1159/000315081[4] CHITYALA P K, WU L, CHOW D S L, GHOSE R. Effects of inflammation on irinotecan pharmacokinetics and development of a best-fit PK model[J]. Chemico-Biological Interactions, 2020, 316: 108 933. [5] ERRIDGE C, BENNETT-GUERRERO E, POXTON I R. Structure and function of lipopolysaccharides[J]. Microbes and Infection, 2002, 4(8): 837 -851 . doi: 10.1016/S1286-4579(02)01604-0[6] ZHOU X, ZHAO R, LV M, XU X, LIU W, LI X, GAO Y, ZHAO Z, ZHANG Z, LI Y, XU R, WAN Q, CUI Y. ACSL4 promotes microglia-mediated neuroinflammation by regulating lipid metabolism and VGLL4 expression[J]. Brain, Behavior, and Immunity, 2023, 109: 331 -343 . doi: 10.1016/j.bbi.2023.02.012[7] ZHOU J, PENG Z, WANG J. Trelagliptin alleviates lipopolysaccharide (LPS)-induced inflammation and oxidative stress in acute lung injury mice[J]. Inflammation, 2021, 44(4): 1 507-1 517. [8] PURIS E, KOUŘIL Š, NAJDEKR L, AURIOLA S, LOPPI S, KORHONEN P, GÓMEZ-BUDIA M, FRICKER G, KANNINEN K M, MALM T, FRIEDECKÝ D, GYNTHER M. Metabolomic, lipidomic and proteomic characterisation of lipopolysaccharide-induced inflammation mouse model[J]. Neuroscience, 2022, 496: 165 -178 . doi: 10.1016/j.neuroscience.2022.05.030[9] WANG G, LIU X, HUO Q, LIN S, WANG W, LIU C, SHA C, LIU W. Optimization and validation of the liquid chromatography coupled to tandem mass spectrometry method for assessing octreotide release from microspheres during inflammation in rabbit models[J]. Journal of Chromatography B, 2023, 1 214: 123 564. [10] AZHAR S, REAVEN E. Scavenger receptor class BI and selective cholesteryl ester uptake: partners in the regulation of steroidogenesis[J]. Molecular and Cellular Endocrinology, 2002, 195(1/2): 1 -26 .[11] GWYNNE J T, STRAUSS J F. The role of lipoproteins in steroidogenesis and cholesterol metabolism in steroidogenic glands[J]. Endocrine Reviews, 1982, 3(3): 299 -329 . doi: 10.1210/edrv-3-3-299[12] SCHIFFER L, BARNARD L, BARANOWSKI E S, GILLIGAN L C, TAYLOR A E, ARLT W, SHACKLETON C H L, STORBECK K H. Human steroid biosynthesis, metabolism and excretion are differentially reflected by serum and urine steroid metabolomes: a comprehensive review[J]. The Journal of Steroid Biochemistry and Molecular Biology, 2019, 194: 105 439. [13] SUN X, ZHANG C, GUO H, CHEN J, TAO Y, WANG F, LIN X, LIU Q, SU L, QIN A. Pregnenolone inhibits osteoclast differentiation and protects against lipopolysaccharide-induced inflammatory bone destruction and ovariectomy-induced bone loss[J]. Frontiers in Pharmacology, 2020, 11: 360 . doi: 10.3389/fphar.2020.00360[14] GHEZZI P, SANTO E D, SACCO S, FODDI C, BARBACCIA M L, MENNINI T. Neurosteroid levels are increased in vivo after LPS treatment and negatively regulate LPS-induced TNF production[J]. European Cytokine Network, 2000, 11(3): 464 -469 .[15] 董俊升. 皮质醇对LPS诱导的BEECs和RAW264.7细胞的炎性损伤调控机制的研究[D]. 扬州: 扬州大学, 2020. [16] NOZAKI M, HARAGUCHI S, MIYAZAKI T, SHIGETA D, KANO N, LEI X F, KIM-KANEYAMA J R, MINAKATA H, MIYAZAKI A, TSUTSUI K. Expression of steroidogenic enzymes and metabolism of steroids in COS-7 cells known as non-steroidogenic cells[J]. Scientific Reports, 2018, 8(1): 2 167. [17] LI Z, JIANG Y, GUENGERICH F P, MA L, LI S, ZHANG W. Engineering cytochrome P450 enzyme systems for biomedical and biotechnological applications[J]. Journal of Biological Chemistry, 2020, 295(3): 833 -849 . doi: 10.1016/S0021-9258(17)49939-X[18] ALMAZROO O A, MIAH M K, VENKATARAMANAN R. Drug metabolism in the liver[J]. Clinics in Liver Disease, 2017, 21(1): 1 -20 . doi: 10.1016/j.cld.2016.08.001 -
计量
- 文章访问数: 521
- HTML全文浏览数: 521
- PDF下载数: 1
- 施引文献: 0